Introduction
As part of the 2021 archaeological excavation campaign of Butrint and Phoenice (Southern Albania) conducted by the University of Bologna and the Institute of Archaeology in Tirana (Figure1),1a new zooarchaeological analysis has been conducted on some of the excavated contexts.2
The faunal remains of the excavations in Phoenice from 2015 to 2019 have been found in sectors A5, A11, A20, A23, C4 and in few tombs in sector A5 too. Moreover, the faunal remains coming from the 2021 excavation in Butrint, from Areas 4, 5, 6, and 7, have been studied as well.3
At the site of Phoenice 1483 osteological remains have been studied in total, and in Butrint 781 remains were analysed. These faunal remains showed a high rate of identification possibly due to some sort of involuntary selection carried by operators while collecting or in some phases of the recovery strategy. For this reason, statistical and quantitative analysis have been limited at this preliminary stage.4
The taphonomic condition of all these faunal remains shows many marks on the surface of the bones, which could be referred both to butchery marks and to gnawing mostly associated to carnivores like dogs. Only very few remains show bite marks by rodents, whereas traces of exposure to burning are completely absent.
Methodology
The zooarchaeological analysis involved a preliminary count of determined osteological remains as well as undetermined ones, filing the resulting data into a specific database file in MS Excel 2019©. When the identification of bones or species was not possible, only the animal size was taken into account. Therefore, these indeterminate remains have been divided into three groups, including vertebrae, ribs and various elements: large, medium and small sized, excessively fragmentary bones for which even original size was not possible to ascertain.
The number of identified species (NISP) has been uploaded on a specific database in MS Access 2019© with additional data, like laterality, preserved part, age of death and sex when available.
After data coming from all the excavated area will be combined on this database, it will allow to obtain results like minimum number of individuals (MNI), as in the case study of Phoenice, although larger samples are necessary to infer more properly on the kill-off patterns economy through every chronological phase of the contexts.
The case study of Butrint allowed to start the zooarchaeological analysis almost simultaneously with the archaeological excavation. As a consequence, the data were filed on the Excel form only, although it has already delivered some information during the excavation. In this case study the faunal remains have not been washed before the preliminary count, allowing to recover more fragments of bones and optimising activities and time during the excavation. Successively, all the determined remains will have to be washed carefully and filed in the Access database, where all data will be elaborated like previously mentioned.
At this initial stage, the anatomic identification of these remains has been carried out by adopting atlases only (Schmid 1972, Barone 1976, Wilkens 2002), since no reference collection was available for consultation at the time. Therefore, in the future some remains will have to be verified, for example through osteometry or biometrical indexes analysis.
Phoenice
The faunal sample of Phoenice shows the level of identification with 703 NISP equal at 47% (Tab. 1, Figure 2). Many of the analysed remains come from the area of the upper town, A11, which is still under excavation (Figure 3). For this reason, the NISP count here should be regarded as preliminary pending a more detailed analysis further on. At the moment, the zooarchaeological analysis brings attention on the other areas, which have been completely excavated and where the fauna has been already analysed.
In Area 23 a basilica belonging to an episcopal complex has been unearthed together with an acceptable amount of faunal remains. The chronological sequence of this area dates back to the 5th century CE when the basilica was built on the Hellenistic ruins, being at last abandoned around the 16th century following multiple construction activities.
Area 5 is the district of the Hellenistic Stoà and a small square church dating to the 7th century CE, which was abandoned in the 16th century CE. Few faunal remains have been found in this area, which can be added to the ones from the tombs in the same area and dating back to the Late Antique and Middle Ages (Cirelli, Podini 2018, 235-247).
Moreover, Area C4 contains a Hellenistic residence named the ‘House of paintings’ dating to 3rd-2nd centuries BCE. Many faunal remains have been found in this house, especially in rooms C2, A1 and D (Gorica 2015, 43-46). Finally, Area 20 represents a small trench excavated within the bath complex of the Late Antique phase, from where the quantity of faunal remains sampled is too small for the zooarchaeological analysis.
As usual, in these samples the domestic species are more abundant. Sheep and goats (Ovis vel Capra) with the pigs (Sus domesticus) are the more common species, followed by cattle (Bos taurus) , whereas there are few remains of equids (Equus sp.), dogs (Canis familiaris) and domestic fowls (Gallus gallus) . In the same sample, there are some wild species probably associated to hunting activities and high numbers of shells suggesting molluscs picking.
In the areas where the excavations were completed, it was possible to estimate the MNI for wild and domestic species. In area A23 the NISP shows a preference for cattle remains followed by pigs and then sheep and goats, while the MNI estimate shows that pigs are more frequent than sheep and goats, as well as cattle.
Pigs are attested by 8 individuals: 2 males older than 4 years old, 2 adults between 2-4 years, with one male and one female, 3 sub-adults between 1-2 years old and a juvenile individual between 4-6 months old.5
Five individuals of sheep and goats were estimated: an adult goat between 4-6 years old, another one between 3-4 years old, 2 sub-adult goats between 1-2 years old and one sub-adult sheep.6
The criteria used to distinguish between sheep and goats follow those suggested in literature.7
Cattle are attested by only 4 individuals: an adult between 4-8 years old, another adult between 3-4 years old and 2 sub-adults between 18-30 months old.8
In area A5 the NISP amounted to a few pieces for each species, while the MNI estimated 4 individuals for pigs, sheep and goats too, and only one cattle. Pigs include: an adult of more than 4 years old, an adult between 2-4 years old, a sub-adult between 12-24 months old and a young between 7-12 months old. The sheep and goats are: a young between 6-12 months old, a sub-adult between 1-2 years old, an adult between 4-6 years old and an adult between 3-4 years old, which could also be a sheep. The identified cattle was an adult between 4-8 years old. This count of MNI stays unaltered, even adding the few remains from the tombs of area A5. These faunal remains are not food offering to the dead, but they are intrusive elements in the soil of the burial, which came from some destruction or reworked contexts.
The estimate of NISP and MNI in area C4 of the Hellenistic house shows more sheep and goats than pigs and cattle. The sheep and goats are 7 individuals: a juvenile goat between 2-6 months old, a sub-adult sheep and a sub-adult goat between 1-2 years old, 2 adult sheep and 2 adult goats, which one between 3-4 years old and another between 2-3 years old. The identified pigs are 5: an adult of more than 4 years old, a female adult between 2-4 years old, 2 sub-adults between 12-24 months old, and a young between 7-12 months old. On the other hand, cattle are 3 individuals: an adult of more than 4 years old, a sub-adult between 18-30 months old and a juvenile between 5-6 months old.
Some preliminary osteometry information allow to have a first estimate of the height at withers:9sheep is tall 65 cm circa based a metacarpal bone, whereas the goat is tall 62 cm approximately on other metacarpal10 ( Tab. 3). Moreover, goats show two types of horns, the first linear and the second twisted, whereas only one horn of sheep was found. The withers height of pigs was estimated between the 66 and 78 cm, based on two metapodials.11
These ones look significantly smaller in size than two others from which it has been estimated a wither’s height between 97 and 104 cm, reasonably associable to wild boars. In addition, the lower canine teeth show a dimensional gap between two species, for example the breadth of root is 2,5 cm in the wild boar and 1,6-1,8 cm in the pigs (Figure 4). Although a proper distinction between wild boar and pig is possible only in adult individuals, whereas it is hard to accomplish as for young individuals or for crossbreeding.
There is not much information for the cattle, because their remains were very fragmented due to butchery. Nevertheless, the measured overall length of an almost complete radio would suggest an individual with withers height of more than 113 cm.12
Moreover, there was a very interesting astragalus with a smooth edge found in room C2 of the ‘House of paintings’ in area C4 (Figure 5).
These objects are quite known in literature and they may have been employed for playing or as payment, but also as religious symbols (Curci 2005,63-65; 2007, 110-117; De Grossi, Minniti 2013; Holmgren 2004). This smoothy astragalus is important because it is the only exemplar found in a residential area, whereas other known astragalus came from the necropolis.
Dog remains are few in the sample and we cannot estimate their physical features or the MNI. Although, there is no trace of these remains inside the ‘House of paintings’ those would be the first ones identified within the residential area of the upper town, because in the previous study they were only attested in the necropolis (Curci 2007, 112-113).
The equid remains are few and always identified as an adult individual, probably a donkey, because the osteometrical data are very similar to the donkey of the tomb no. 27 of the necropolis (Curci 2005, 109-112).
In the archaeological sample, NISP count shows that between the wild species more frequently occur are the red deer ( Cervus elaphus ) then the wild boar ( Sus scrofa ) . The deer prefers living in woods with clearings, whereas the wild boar can live in the bush very well. About the deer remains, there are no fragments of antlers but only metapodials and a few fragments of radius, scapula, tibia and femur. All these remains show a lot of butchery marks: the occurrence of a metapodial and phalanges can prove the existence of a butchery area nearby, because these remains have little alimentary relevance (Figure 6). The estimate of the MNI of these wild species of area A23 shows an adult deer individual older than 3 and half years old, based on the fused epiphysis of distal radius,13 and a wild boar of more than 4 years old, estimated by lower canine tooth. In area C4 the presence of two deer have been estimated on the basis of two left astragali, whereas the remains of wild boar belonging to an adult individual of more than 2 years old was estimated on two metacarpals.14
The few remains of the fallow deer ( Dama dama ) and the roe deer ( Capreolus capreolus ) have also been preliminarily identified: to the former would belong a phalange and one fragment of metacarpal, whereas the roe deer was identified by a fragment of distal tibia and two scapulas. The fallow deer lives in an environment of woods with clearing, whereas the roe deer can adapt to live in the bush very well and near areas occupied by humans.
The distal radius of the fox ( Vulpes vulpes ) will require a further check with a reference collection, because it is the first time the species has been identified at the site of Phoenice. In addition, some fragments of radius, scapula, and tibia have been attributed to an indistinguishable rabbit or hare (Leporidae). Moreover, fragments of badger ( Meles meles ) found in the ‘House of two peristyle’ will need too more verification. A few elements belonging to reptiles have also been determined, assigned in particular to few remains of pond ( Emys sp.) and land tortoise ( Testudo sp.).15
As regards birds, many remains have been attributed to domestic fowls. Though a small coracoid was provisionally attributed to a wild species of the galliformes order, by morphology and size would look compatible with a rock partridge ( Alectoris graeca ). Instead, a large-sized wing phalange has furthermore been collected but not yet determined.
The archaeomalacology analysis has been important for this case study, because at the moment it is the only information available about the marine resources.16
The identified species and family are as follows:17
Marine Bivalve
• Cerastoderma sp.*
• Glycymeris sp.
• Mytilus sp.
• Ostrea edulis*
• Pecten jacobaeus
• Spondylus gaederopus
•Veneridae*
Marine Gastropod
• Bolinus brandaris
• Hexaplex trunculus*
• Tarantinaea lignaria
Terrestrial Gastropod
•Helicidae*
• Pomatias elegans
The main species found in the site used as food is the common cockle ( Cerastoderma sp.) followed by the thorny oysters (Spondylus gaederopus), which has a big relevance for the alimentation, since it tastes like oysters18 (Figure 7) There were not worked pieces of these shells on the site, therefore this shell was not used as ornament but only as food. Moreover, these molluscs were picked by diving down to a depth between 2-30 meters on the rocky seabed.
Belonging to the Muricidae family are attested the murex trunculus (Hexaplex trunculus) and the spiny dye-murex ( Bolinus brandaris ), both commonly associated to the dyeing process. The murexes were consumed also for food, so its presence does not automatically suggest purple dye extraction. Moreover, the structures for smashing and macerate these molluscs have not been found in Phoenice yet. Another marine gastropod found in the site was a spindle snail ( Tarantinaea lignaria ), whereas among the marine bivalves there were some species of the Veneridae family, flat oysters ( Ostrea edulis ) , mussels ( Mytilus sp.) , a fragment of saltwater clam ( Glycymeris sp.) and other fragment of scallop ( Pecten jacobaeus ).19
The few terrestrial gastropods found in the excavated layers were supposed to be intrusive during the process of natural accumulation. Nevertheless, these types of snails of the Helicidae family are edible, whereas the specie Pomatias elegans is not edible and describe an arid environment with dry leaves on the ground (Wilkens 2002, Girod 2015).
Butrint
The faunal sample of the Butrint shows the level of identification with 452 NISP equal at 57% (Tab. 2, Figure 8). These remains come from the layers excavated during 2021, located on the central plateau of the Acropolis area near the western perimetral wall, which has different phases of construction, going from the Archaic to the Medieval period. The Italian-Albanian mission has excavated in this area since 2019. During the last campaign, two trenches have been opened in the filling ( emplekton ) of the polygonal Archaic wall and two other few meters to the North, where some medieval structures built above a Roman complex were found (Giorgi 2018; 2022; Giorgi, Lepore 2020; Giorgi, Lepore, Gamberini 2020). The analysed faunal remains come from these four investigated areas. The first two areas to the North are dated to the Medieval and to the Roman-Hellenistic period, while those ones near the polygonal wall are dated to the Archaic period (Figure 9).
At a first glance these faunal remains seem to come from layers of collapse or abandonment containing heterogeneous materials. In this sample, the highest NISP came from Areas 4 and 5. On the other hand, only few faunal remains were found in Areas 6 and 7, since previous excavations had exposed the upper layers of this stratigraphic sequence.
A preliminary chronological framework of faunal remains within Area 4 has been dated to the Roman period. In the so-called Room VI20 located in this same area, many faunal remains come from the Roman layers, while a small quantity was found in SU 434 dating to the Hellenistic period provisionally. In Area 5 the faunal remains come from the Medieval layers, except for a few ones, which have been excluded since they have been collected in upper and rather unreliable contexts. While a lot of shells have been found within preserved Archaic layers of Areas 6 and 7 mainly.21
This sample shows that the NISP of pig is the most attested, followed by sheep and goats, and then cattle. Regarding other domestic species just one element could be assigned to dog, whereas the equid remains were not found yet.
At Butrint there were wild animals like red deer, wild boar, and hare. In the sample no traces of deer antlers have been recognized, but only its osteological remains. On the other hand the remains of wild boar were found in stratigraphy of Area 4 and Room VI. In the latter, it was also found a fragment of pelvis belonging to a lagomorph, but its poor condition does not allow to distinguish if it belonged to a rabbit or a hare. Among the birds found in the sample, there are some remains of domestic fowls, but in the Archaic stratigraphy of perimetral wall a small humerus was found also, which has been tentatively attributed to a no specified passerine ( Passeriformes ). Currently, as for the previous case study of Phoenice, information about fishing is not available yet due to the difficulty in recovering these small and fragile remains. However, a sample of soil collected in Room VI located in Area 4, even though not studied yet, has showed the presence of some fish and micromammal remains.22
In addition, in the superficial layer of Area 5 the caudal spine of a ray-like species ( Rajiformes ) has been preliminarily identified.
To sum up, all these preliminary data still have to be checked against through MNI and others analyses. Moreover, the sample needs a bigger quantitative of faunal remains to submit reliable statistical data about the chronological sequence of the site.
In this case study, the archaeomalacology analysis allowed to gather important data on the exploitation of water sources (Figure 10). A lot of these shells have been unearthed in a dumping fill disposed inside room VI in area 4 during the abandonment process. The great number of shells found among the stone-rich infill of the emplekton are the main source of faunal information for the Archaic period, since their structure composed by calcium carbonate allows a better preservation than other organic remains. The identified species and family are:
Marine Bivalve
• Cerastoderma sp.*
•Veneridae*
• Ostrea edulis
• Mytilus sp. *
• Spondylus gaederopus*
Marine Gastropod
• Bolinus brandaris
• Hexaplex trunculus*
• Patella sp.
• Cerithium sp.*
• Monodonta sp.*
Terrestrial Gastropod
•Helicidae
• Pioretia sp.
Freshwater Bivalve
• Unio sp.
The presence of the same species found at Phoenice would suggest a similar activity of marine shell picking taking place at both sites, where their supply areas had the same environmental features. The exception are the ceriths snail ( Cerithium sp.) and the toothed topshell ( Monodonta sp.), which have not been found at Phoenice, but they had already been found during the Forum excavation (Hernandez 2017, 223-224). The toothed topshell prefers rocky seashore, where it can hide among the meadows of Posidonia oceanica . Instead, the ceriths snail prefers low rocky seashore or sandy seabed with a lot of detritus. Their spiral shape can suggest an ornamental use, but these molluscs have also a small alimentary purpose.
For what concerns land snails, some individuals of the Helicidae family, which prefer wet and shaded habitats, were identified in the site. These terrestrial gastropods are edible, but their occurrence at the site could also be due to a natural intrusion during the accumulation phases of the layer. On the other hand, other land snail species (Poiretia sp.)23are not edible, hinting at the surrounding environmental features during the accumulation phases of the layer, consisting of wooded environment with dead leaves on the ground. However, this snail has also been collected in the oldest phases near the perimetral wall and in Area 4. Finally, in the upper layer of Area 6 a freshwater mussel (Unio sp.)24was found for the first time. This mollusc lives in ponds or weak flowing rivers with muddy riverbed. In antiquity they were used for alimentary purpose and for craftworks exploiting its internal mother-of-pearl surface (Wilkens 2002; Girod 2015).
Final considerations
This zooarchaeological analysis relative to the 2021 archaeological campaign intends to offer a preliminary glance to the animal exploitation in the investigated Epirote area: more detailed analysis and larger samples will be of course necessary to piece together a more accurate picture on its role in the local economy.
These two sites of Chaonia region have relied on apparently very similar economies, where the breeding of animals, like pigs, cattle, sheep and goats, seems to have been the chief practice to obtain animal-based resources. Moreover, according to the MNI estimated from the site of Phoinike, it is possible to suppose that pig represented the more frequently exploited domestic species during Late Antiquity. On the contrary, in the older periods sheep and goats were preferred than other domestic species.
Beside breeding, hunting had also a substantial importance at these sites, being practiced on large and small extensive woodlands and clearings. For what concerns the birds, domestic fowls were the most common, while few wild species were hunted infrequently. Such frequencies look pretty consistent with those collected in the Thesprotia district (Niskanen 2009, 145-154; Deckwirth 2011, 297-309).
On the other hand, we do not have any information about fishing, which was probably very influential at the Butrint site thanks to its strategic position at close range to sea, river and lagoon. However, some preliminary information came from the archaeomalacological analysis. These shells describe two different kind of water sources and environments, and also two different modalities of mollusc picking practice:
•The former is a marine environment represented by a rocky intertidal zone between low and medium depth, featuring meadows of Posidonia oceanica , where target of shells collection might have been species like: Hexaplex trunculus , Spondylusgaederopus, Cerithium genus Poiretia,Monodonta genus Poiretia and Patella genus Poiretia.
•The second type is a lagoon environment, with sandy or muddy seabed, in such context the mainly picked species isCerastoderma Posidonia oceanica, which might have been the primary mollusc for food purpose . This species populates the lagoon environment abundantly and they can be picked up during low tide, as well as with trawling nets. In this habitat it might occur also the Mytilus sp. and the Ostrea edulis , which prefer water with low salinity. Since there is no trace indicating they might have been the result of farming, they seem to have been picked up fortuitously, even if it depends largely on the quantitative samples studied.25
Finally, some rare attestation of species like: Bolinus brandaris, Pecten jacobaeus, Glycymeris sp. and Veneridae family, which prefer a sandy and soft seabed, therefore they can come from a small environment of Epirus or a different seashore.
In conclusion, the amount of osteological remains investigated at the two sites is rather different, making an accurate comparison between the two faunal samples difficult. However, data appear encouraging and, in the future, they will be integrated by useful information gathered from soil samples, which will increase the quality of the data and improve the statistical power of the survey.
Bibliography
- Barone, R. 1976, Anatomie comparée des Mammiféres domestiques , Osteologia, Bologna: Edagricole.
- Böessneck, J., H.H. Müller, M. Teichert. 1964, ‘Osteologische Unterscheidungsmerkmale zwischen Schaf (Ovis aries) und Ziege (Capra hircus)’, Kühn Archiv , 78, 1-129.
- Böessneck, J. 1969, ‘Osteological differences between sheep (Ovis aries Linné) and goat (Capra hircus Linné)’, in Science in Archaeology. A Survey of Progress and Research , edited by D. Brothwell, E. and Higgs, 331-358, London: Thames & Hudson.
- Bull, G., Payne, S., 1982, ‘Tooth eruption and epiphisial fusion in pigs and wild boar ’ , in Ageing and sexing animal bones from archaeological sites , edited by B. Wilson, C. Grigson, S. Payne, 55-72, Oxford: B.A.R.
- Cirelli E., Podini M. 2018, ‘Gli edifici ecclesiastici di Phoinike e del suo territorio tra tarda Antichità e Medioevo ’,Hortus Artium Mediaevalium , Journal of the International Research Center for Late Antiquity and Middle Ages, 24, 235-247.
- Curci, A. 2005, ‘Dati sull’alimentazione dall’analisi dei resti faunistici’ In Phoinike III Rapporto preliminare sulle campagne di scavi e ricerche 2002-2003, edited by S. De Maria, S. Gjongecaj, 63-66, Bologna: Ante Quem.
- Curci, A. 2005, ‘Il funerale dell’asino e la tomba 27’, In Phoinike III Rapporto preliminare sulle campagne di scavi e ricerche 2002-2003, edited by S. De Maria, S. Gjongecaj, 107-112, Bologna: Ante Quem.
- Curci, A. 2007, ‘Osservazioni sui resti faunistici ’ , In Phoinike IV Rapporto preliminare sulle campagne di scavi e ricerche 2004-2006,edited by S. De Maria, S. Gjongecaj, 110-117, Bologna: Ante Quem.
- Deckwirth, V. 2011, ‘A tower of meals: Trenches A and F of Agios Donatos’, In Thesprotia Expedition II: Environment and Settlement Patterns, edited by B. Forsén, 297-309, Helsinki: Finnish Institute at Athens.
- De Grossi Mazzorin, J. 2015, ‘Consumo e allevamento di ostriche e di mitili in epoca classica e medievale ’ , in Appunti di Archeomalacologia, edited by A. Girod, 153-158, Firenze: All’insegna del Giglio.
- De Grossi Mazzorin, J., Minniti C. 2013, ‘Ancient use of the knuckle-bone for rituals and gaming piece’, Anthropozoologica, 48, 371-380.
- Giorgi, E. 2017, ‘Landscape and Citizens during the early Roman era in Northern Epirus: Phoinike and the Chaonia region (2nd BC-2nd AD)’, Groma, 2, 1-20. DOI: https://dx.doi.org/10.12977/groma9
- Giorgi, E. 2018, ‘The Destruction of Epirus after Pidna. Archeology and Literature’,Dialoghi di Archeologia della Magna Grecia e del Mediterraneo, edited by M. Cipriani, M. Scafuro, and A. Pontrandolfo, 91-105. Paestum: Pandemos.
- Giorgi, E. 2022. ‘Another Greece. The Population of Northern Epirus and the Genesis of Butrint in the Archaic Period.’ In Schemata. La città oltre la forma. Per una nuova definizione dei paesaggi urbani e delle loro funzioni: urbanizzazione e società nel mediterraneo pre-classico. Età arcaica, edited by R. Brancato, L.M. Caliò, M. Figuera, G.M. Gerogiannis, E. Papalardo, and S. Todaro, 477–502. Cronache - Monografie. Roma: Edizioni Quasar.
- Giorgi, E., Lepore, G., 2020, ‘Comparing Phoinike and Butrint. Some remarks on the walls of two settlements in Northern Epirus ’ , in Fortificazioni e società nel Mediterraneo occidentale , edited by L. M. Caliò, G. M. Gerogiannis, M. Kopsacheili, Roma: Edizioni Quasar.
- Giorgi, E., Lepore, G., Gamberini, A. 2020, Boundaries Archaeology: Economy, Sacred Places, Cultural Influences in the Ionian and Adriatic Areas,Heidelberg: Propylaeum.
- Girod, A.,2015 , Appunti di Archeomalacologia , Firenze: All’insegna del Giglio.
- Gorica, S. 2015, ‘La casa dei dipinti’, In Phoinike VI Rapporto preliminare sulle campagne di scavi e ricerche 2011-2014, edited by S. De Maria, S. Gjongecaj, 43-49, Bologna: Ante Quem.
- Grant, A. 1982, ‘The use of tooth wear as a guide to the age of domestic ungulates’, in Ageing and sexing animal bones from archaeological sites , edited by B. Wilson, C. Grigson, S. Payne, 91-108, Oxford: B.A.R.
- Greenslade, S., Leppard S., Logue M. 2012, ‘The acropolis of Butrint reassessed’ in Butrint 4: The archaeology and histories of an Ionian Town , edited by I.I Hansen, R. Hodges, and S. Leppard. Oxford: Oxbow Books, 47-77.
- Hernandez, D. R. 2017, ‘Bouthrotos (Butrint) in the Archaic and Classical Periods: The Acropolis and Temple of Athena Polias’, Hesperia , 86, 205-271.
- Holmgren, R. 2004, ‘Money on the hoof: The astragalus bone religion, gaming and primitive money’ in Pecus: Man and Animal in Antiquity , edited by Frizell B. F., 212-220. Proceedings of the conference at the Swedish Institute in Rome, September 9-12, 2002, Rome: The Swedish Institute.
- Mariezkurrena, K. 1983, ‘Contribución al conocimiento del desarrollo de la dentición y el esqueleto postcranial de Cervus elaphus’, Munibe Antropologia Arkeologia, 35, 149-202.
- Matolcsi, J. 1970, ‘Historische Erforschung der Körpergröbe des Rindes auf Grund von ungarischem Knochenmaterial’, in Zeitschrift für Tierzüchtung und Züchtungsbiologie , 87, 89-137.
- Micheli, R. 2015, ‘Conchiglie affascinanti per ornamenti di prestigio: produzione, distribuzione e impiego dello Spondylus gaederopus nella preistoria europea’, in Appunti di Archeomalacologia, edited by A. Girod , 198-210, Firenze: All’insegna del Giglio.
- Niskanen, M. 2009, ‘A shift in Animal species used for food from early Iron Age to roman period’, in Thesprotia Expedition I: Towards a Regional History , edited by B. Forsén, 145-154, Helsinki: Finnish Institute at Athens.
- Payne, S. 1973, ‘Kill-off Patterns in Sheep and Goats: The mandibles from Asvan Kale ’ , in Anatolian Studies , 33: 65-81.
- Prummel, W., Frisch, H.J. 1986, ‘A guide for the distinction of species sex and body side in bones of sheep and goat’, Journal of Archaeological Science , 13: 567-577.
- Schmid, E. 1972, Atlas of animal bones. For Prehistorians, Archaeologist and Quaternary Geologists, Amsterdam, London, New York: Elsevier Publishing Company.
- Schramm, Z. 1967, ‘Long bones and height in withers of goat’, Roczniki Wvzszej Szkolv Rolniczei w Poznaniu , 36: 89-105.
- Silver, I. A. 1969, ‘The ageing of domestic animals’, in Science in Archaeology, edited by D.R. Brothwell, E.S. Higgs, 283-302, London: Thames & Hudson.
- Teichert, M. 1969, ‘Osteometrische Untersuchungen zur Berechnungen del Widderristhöhe bei vor-und
- frühgeschictlichen Schweinen’, Kühn Archiv 83: 237-292.
- Von Den Driesch, A. 1976, A guide to the measurement of animal bones from archaeological sites, Peabody Museum Bulletins 1, Cambridge: Harvard University.
- Welter-Schultes, F. W. 2012, European non-marine molluscs, a guide for species identification, Gottingen: Planet poster editions.
- Wilkens, B. 2002, Archeozoologia. Manuale per lo studio dei resti faunistici di area mediterranea, Dipartimento di Storia Università degli Studi di Sassari, CD-Rom.
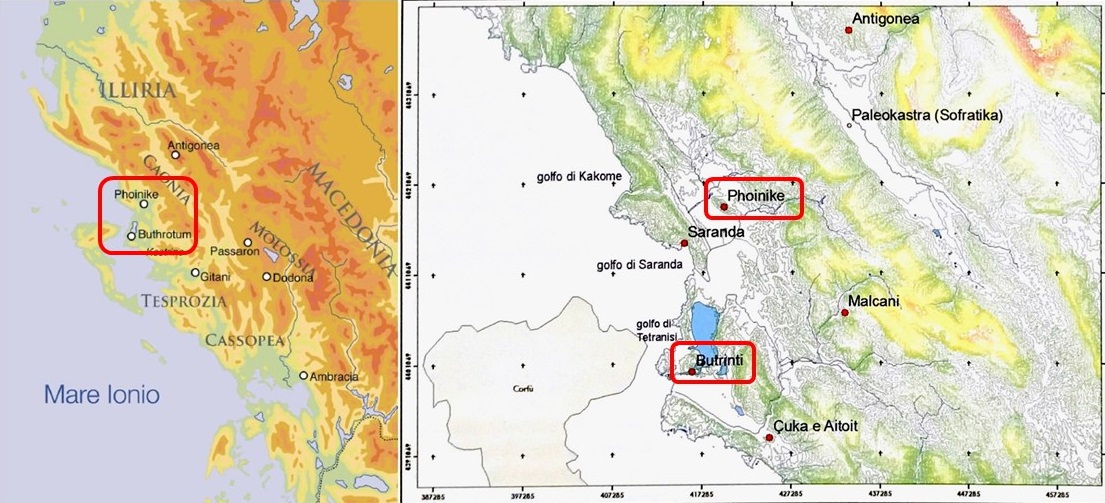 Figure 1. Location of the sites of Phoenice and Butrint. After Giorgi 2017, 2-3, Figure1-2.
Figure 1. Location of the sites of Phoenice and Butrint. After Giorgi 2017, 2-3, Figure1-2.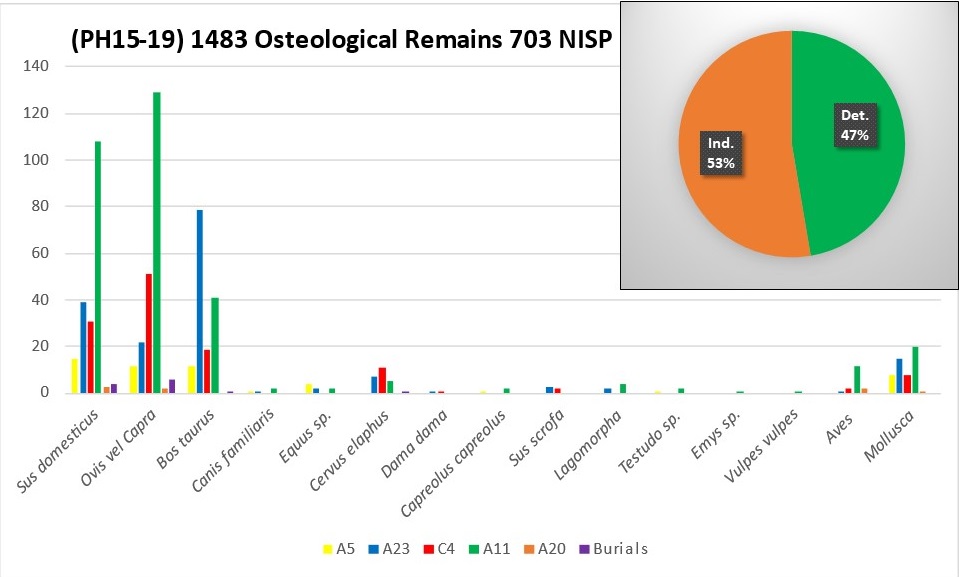 Figure 2. Pie chart with the rate of identification (%) of all the osteological remains, and bar chart of NISP sorted by excavated areas during 2015-2019 archaeological seasons (PH15-PH19)
Figure 2. Pie chart with the rate of identification (%) of all the osteological remains, and bar chart of NISP sorted by excavated areas during 2015-2019 archaeological seasons (PH15-PH19)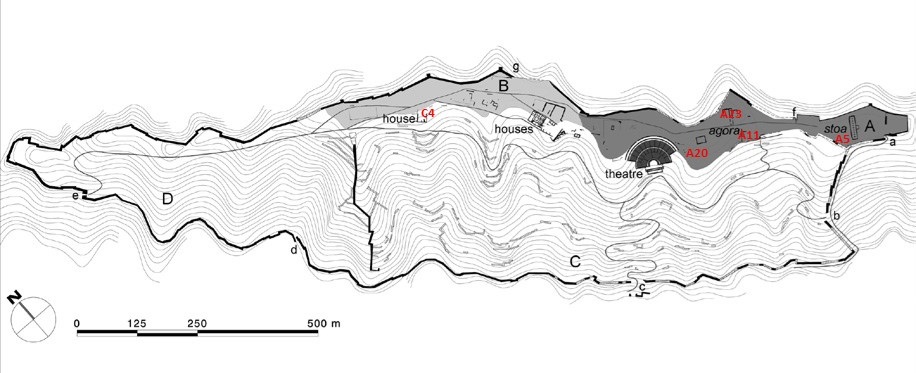 Figure 3. Map of Phoenice with analysed areas. After Giorgi, Lepore 2020, 155, Figure 2.
Figure 3. Map of Phoenice with analysed areas. After Giorgi, Lepore 2020, 155, Figure 2. Figure 4. Difference in Lower male canine teeth of wild boar and pigs.
Figure 4. Difference in Lower male canine teeth of wild boar and pigs. Figure 5. Smoothed cattle astragalus.
Figure 5. Smoothed cattle astragalus. Figure 6. the second phalanx of a red deer with cut-marks.
Figure 6. the second phalanx of a red deer with cut-marks.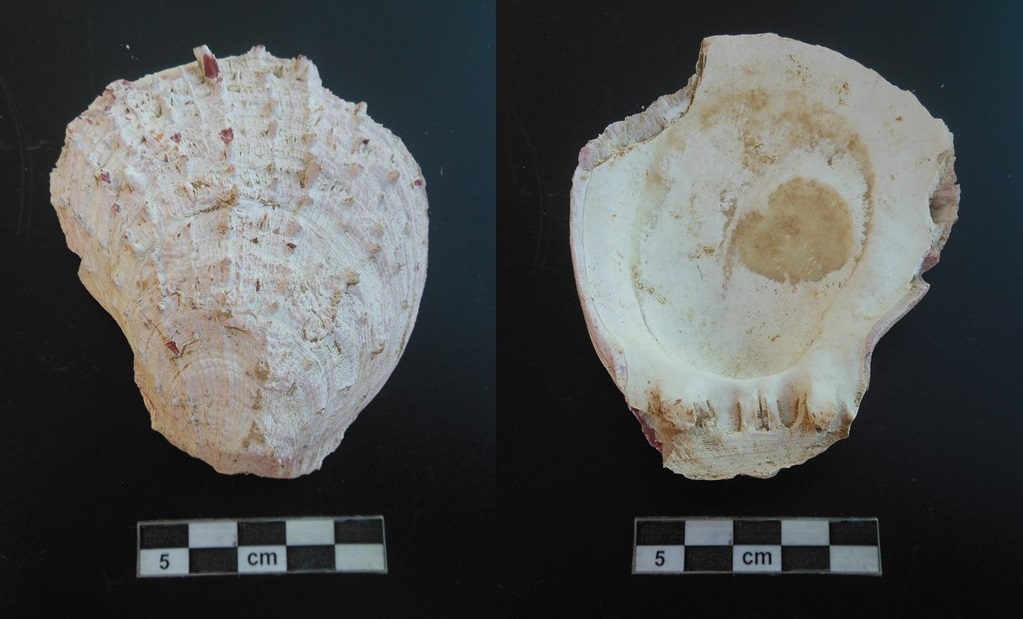 Figure 7. Specimen of Spondylus gaederopus found in the site of Phoenice.
Figure 7. Specimen of Spondylus gaederopus found in the site of Phoenice. Figure 8. Pie chart with the rate of identification (%) of all the osteological remains, and bar chart of NISP for excavated areas during 2021 archaeological season (BU21).
Figure 8. Pie chart with the rate of identification (%) of all the osteological remains, and bar chart of NISP for excavated areas during 2021 archaeological season (BU21).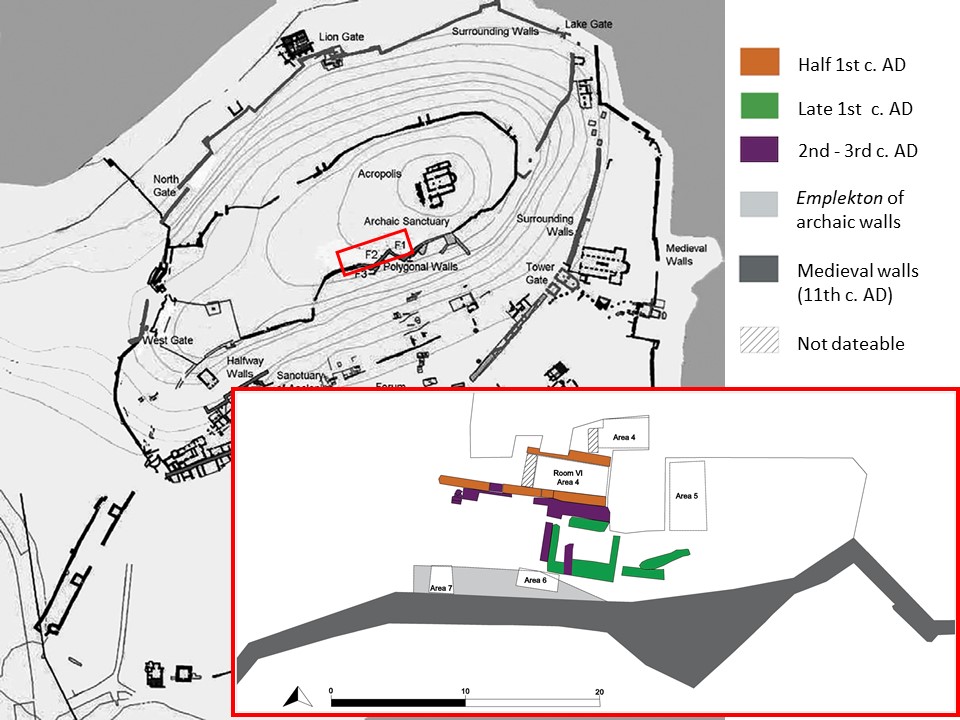 Figure 9. Map of Butrint with analysed areas, elaborated by Federica Carbotti. The colors are same used at the map by Butrint Foundation (Greenslade Greenslade, Leppard, Logue 2013).
Figure 9. Map of Butrint with analysed areas, elaborated by Federica Carbotti. The colors are same used at the map by Butrint Foundation (Greenslade Greenslade, Leppard, Logue 2013). Figure 10. Some sea shells from Butrint.
Figure 10. Some sea shells from Butrint.